A. N. Leontyev 1981
The evolution of the Psyche in Animals
Source: Paul Ballantyne's site at York University;
Edited extracts from Chapter 2: Leontyev, A.N. (1981). “An outline of the evolution of the psyche” (pp. 156-326).
Problems of the Development of the Mind. (Trans. M. Kopylova) Moscow: Progress Publishers.
An outline of the evolution of the psyche
[Section] I. The evolution of the Psyche in Animals
1. [The Stage of Elementary Sensory Psyche]
The origin of sensitive living organisms is associated with complication of their vital activity. This complication consists in a differentiation of the processes of external activity that mediate organism's relations with those properties of the environment on which... their life depend.
The differentiation of these processes is due to the [evolutionary] development of [a specialized] irritability to effects that perform a signaling function [i.e., sensitivity]. So a capacity arises for organism's reflection of the effects of surrounding reality in their objective connections and relations, i.e. psychic reflection.
These forms of psychic reflection are developed along with complication of the organism's structure and depending on the development of the activity together with which they originate. Scientific analysis of them is therefore impossible other than on the basis of a survey of the activity of animals itself.
What then is the activity of animals with which the simplest form of their psyche is associated? Its main feature is that it is induced by some property or another affecting the animal to which it is at the same time directed, but which does not coincide with the properties that the animal's life directly depends on. It is governed, consequently, not by the affecting properties in themselves but rather by them in their relation with other properties.
We know, for example, that as soon as an insect gets caught in a spider-web, the spider immediately moves toward it and begins to enmesh it with... thread. What is it that causes this activity of the spider's, and to what is it directed? ... [p. 157]... It has been established experimentally... that what stimulates the spider's activity, and that to which it is directed, is the vibration produced by the wings of the insect... transmitted along the spider-web. As soon as the vibration ceases, the spider stops moving toward its victim. It is enough, however, for the insect's wings to begin vibrating again, for the spider to move toward it again and once more enmesh it in its web. Is it, however, in fact the vibration that evokes the spider's activity and at... what [evolutionary end is] this activity... directed? This is demonstrated by the following experiment. A resonating tuning fork is attached to the spider-web. In response the spider runs to the tuning fork, climbs on to it, spins a web around it, and tries to strike it with its mandibles (Rabaud [(1924)]). This means that it is a matter here of the fact of vibration; for, apart from vibration, there is nothing in common between a tuning fork and an insect that has been trapped in a spider-web.
Why is the spider's [sensory-motor] activity associated precisely with the effect on it of vibration which in itself, of course, plays no [directly assimilative] role in its life? Because, in normal conditions, the effect of vibration has a certain association, a certain stable relation with the nutrient matter of the insect that has been caught in the web. We shall call this relation of an influencing property to satisfaction of a biological need the biological sense of this influence. Employing this term we can say that the spider's activity is directed to a vibrating body because the vibration has acquired a sense of food for it in the course of the species' evolution.
The biological sense of any influence is not constant for an animal, but on the contrary varies and develops during its activity in accordance with the objective associations of the corresponding properties of the environment.
If, for example, a hungry toad is first systematically fed with worms and then an ordinary matchstick and round bit of skin [are] put in front of it, it will pounce on the match stick, which has an elongated form like worms, and not touch the skin; elongated form has acquired the biological sense [p. 158] of food for it. If, on the contrary, we feed the toad first with spiders, it will then pounce on the skin [or on a piece of moss], similar in shape to a spider, without touching the matchstick; the round form of the object has now acquired the sense of food for it.
The sense connections that arise in the activity of animals, it must be noted, are conditioned reflexes with a specific, and (one can even say) extraordinary character. They differ markedly from the conditioned associations that form the mechanism of behaviour itself, i.e., connection by which behaviour is realised.
When an animal, seeing food, moves toward it, i.e. when we are concerned with the sense association 'sight of food- food', this association arises and changes quite differently from those that arise in it, for example, during the formation of a [bodily] habit of avoiding obstacles in its path (an association 'obstacle-by-passing movement').
Links of the first kind [(even at the stage of elementary sensory psyche)] are formed very quickly, as research has shown, from 'scratch', and are broken down just as quickly; one or two combinations are sufficient for that.
Links of the second kind arise and fade slowly, on the contrary, and gradually. Chicks, for example, already begin to peck selectively at chopped egg yolk after a single success. For two-day-old chicks it is sufficient to peck once or twice at a bit of bitter orange peel instead of egg yolk for its food behaviour toward yolk to be wiped out (Morgan, et al). On the other hand, it takes dozens of trials to develop a quite satisfactory adoption of pecking movements in chicks to the external conditions in which they are given food.
Buytendijk [(1930)], studying the forming of habits in toads, gave one of these animals, in one of a series of his experiments, insects whose substance caused a sharp negative biological reaction. A single experience was enough for the toad to refuse for many hours afterward to try and eat these insects, or any other outwardly resembling them. In other experiments he separated the bait (an earthworm) from the toad by glass; in those conditions, the toad on the contrary, displayed great persistence, in spite of its striking the glass each time; it made many attempts before its reaction faded. Even intensifying the moment of 'punishment' [p. 159] (negative reinforcement) did not stop movements in such cases. In Abbot's experiments the frog continued for 72 hours to pounce on bait surrounded by needles, until the skin of its upper jaw was seriously lacerated. The biological significance of the difference in rate of formation of both kinds of association is quite understandable if we allow for the species' life conditions. If a toad (Buytendijk says) approaches an anthill during its evening hunt and catches a poisonous ant, the rapid formation of an association protects it from swallowing other such insects harmful because of the acid they have. When a toad, on the contrary, tries to catch an earthworm but does not succeed in doing so, a second attempt may help it all the same, in ordinary circumstances, to get food.
Another feature of [such] sense connections is their sort of 'bilateral' character, which is expressed not only in the effect of a given stimulus beginning to evoke a certain reaction... [or] ... behaviour, as a result..., but also in a corresponding need now [for the organism] to 'recognize itself' as it were in the object-stimulus, ... and to evoke active, search activity in relation to it.
Darwin [1885] had already stressed the special nature of these sense links, in citing, for example, the following observations:
Thus it is asserted that if a calf or infant has never sucked its mother, it is very much easier to bring it up by hand than if it has sucked only once. So again Kirby states that larvae after having 'fed a time on one plant, will die rather than eat another, which would have been perfectly acceptable to them if accustomed to it from the first' [Posthumous essay on instinct, in Romanes, 1885].
In the classic works of Pavlov and his associates, the formation of such 'rapid' sense associations (in the early work of Tsitovich, and later in the experiments of Narbutovich and others) was also demonstrated, although their special role in behaviour was not... stressed.
An animal's reflection of its environment forms a unity with its [externally observable] activity, which means that, although there is a difference between them, they are at the same time inseparable [p. 160] from one another. This means, furthermore, that there are reciprocal transfers between them, ... ; [in any particular (naturalistic or experimental) case] whether the object's property affecting [an individual] animal is reflected in its sensations, and how exactly, thus depends on whether the animal is really linked with the object in the course of its adaptation to its environment and how precisely it is linked with it. On the other hand, any activity of an animal's mediated by an influence sensed by it is performed in accordance with how the given influence is reflected in its sensations. The animal's activity, that links it in practice with objective reality, is understandably basic in this complex unity of reflection and activity; psychic reflection of the affective property of this reality is secondary and derivative.
The activity of animals in the earliest, first stage of the evolution of the psyche is characterised by its corresponding to some one separate... property (or aggregate of separate properties) by virtue of the property's essential connection with the influences on which performance of their basic biological functions depends. The reflection of reality connected with this structure of activity correspondingly has the form of sensitivity to separate affective properties..., the form of elementary sensation. We shall call this stage in the evolution of the psyche the elementary sensory psyche. It covers a long series of animals, and it is possible that certain higher infusoria possess elementary sensitivity.
We can say that with much greater confidence in relation to such animals as certain worms, crustaceans, insects, and of course in relation to all [lower] vertebrates [e.g., fish].
Variability of behaviour in consequence of established new connections has been demonstrated in worms by many researchers. Annelids, for example, as the experiments of Copeland and Brown [(1934)] have shown, either do not react at all to being touched by a glass rod, or react negatively. When, however, the touch of the rod is associated with feeding, then the worm's reaction alters: touching now evokes a positive reaction to food in it.
Changes of this kind can acquire a more complicated [p. 161] character in crustaceans. When, for example, the abdominal part of a hermit crab is lightly stimulated mechanically when it is in a shell, that will evoke certain movements in it, as Ten Cate-Kazejewa's [(1934)] work has shown. If, however, the stimulation is continued, the animal will quit the shell and run away. That fact is of little interest in itself; what is interesting is the crab's subsequent behaviour. If the experiment is systematically repeated, it turns out, the animal's behaviour becomes different. Now it removes its abdomen from the shell at the first touch, but never deserts it and almost immediately takes up its former position. The touch has now acquired a quite different meaning -it has become a signal for withdrawing the abdomen from the shell.
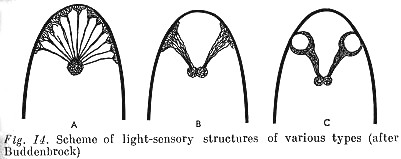 The material basis for the development of animal's activity and sensitivity is understandably their anatomical organization. The common pattern of changes in organisms with which evolution at the stage of elementary sensory psyche is linked, consists on the one hand in animal's organs of sensitivity becoming more and more differentiated at that stage of evolution, and their numbers increasing; correspondingly their sensations are also differentiated. Cells irritable in relation to light in lower animals, are scattered over the whole surface of the body so that these animals can have only very diffuse light sensitivity.
The material basis for the development of animal's activity and sensitivity is understandably their anatomical organization. The common pattern of changes in organisms with which evolution at the stage of elementary sensory psyche is linked, consists on the one hand in animal's organs of sensitivity becoming more and more differentiated at that stage of evolution, and their numbers increasing; correspondingly their sensations are also differentiated. Cells irritable in relation to light in lower animals, are scattered over the whole surface of the body so that these animals can have only very diffuse light sensitivity.
Light-sensitive cells are gathered together at the anterior end of the body later, for the first time, in worms (Fig. 14, A), and, on becoming concentrated acquire the form of plates (B); these organs already provide a possibility [p. 162] of quite precise orientation in the direction of light. Finally, at an even higher stage of evolution (mollusks), an internal, spherical light-sensitive cavity arises through bending of these plates, that operates like a camera lucida (C) that enables the movement of objects to be [sensed].
 On the other hand organs of locomotion are also evolved, organs of animal's external motion. Their evolution occurs specially noticeably in connection with the following two main alterations: on the one hand in connection with the transition to life in a terrestrial environment, and on the other hand, in hydrobionts (animals living in an aquatic environment) in connection with transition to active pursuit of prey.
On the other hand organs of locomotion are also evolved, organs of animal's external motion. Their evolution occurs specially noticeably in connection with the following two main alterations: on the one hand in connection with the transition to life in a terrestrial environment, and on the other hand, in hydrobionts (animals living in an aquatic environment) in connection with transition to active pursuit of prey.
Along with development of organs of sensitivity and organs of locomotion, an organ of connection and coordination of processes -a nervous system- also evolves.
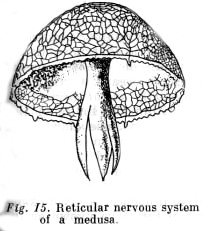
Originally the nervous system is a simple network the fibres of which running in various directions unite sensitive cells located on the surface directly with the animal's contractile tissue. This type of nervous system is not found in contemporary species. In jellyfish or medusae the nerve net leading from sensory cells is connected with muscular tissue already by means of motor nerve cells.
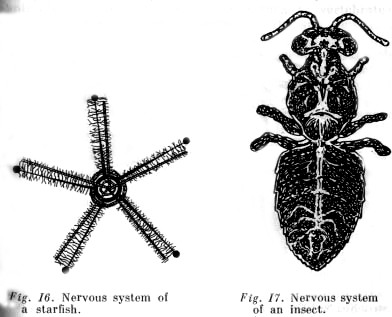 In such a reticular nervous system excitation is diffusely transmitted; the nerve fibres forming the network have two-directional conductivity, and inhibitory processes do not apparently exist. The next step in the evolution of the nerve system is represented in the differentiation of neurons that form central ganglia (nerve junctions). Along one line of evolution (among echinoderms) the nerve ganglia form an apipharangeal ring with nerve columns running off from it. This is already a nerve centre that enables relatively very complexly coordinated movements to be performed, such as a starfish's movements to open the shells of bivalves. Along two other major lines of evolution (from the primitive worms to crustraceans and spiders, and from primitive worms to insects) a more massive anterior... ganglion becomes formed which subordinates the working of lower-lying ganglia to itself (see Figs. 15-17).
In such a reticular nervous system excitation is diffusely transmitted; the nerve fibres forming the network have two-directional conductivity, and inhibitory processes do not apparently exist. The next step in the evolution of the nerve system is represented in the differentiation of neurons that form central ganglia (nerve junctions). Along one line of evolution (among echinoderms) the nerve ganglia form an apipharangeal ring with nerve columns running off from it. This is already a nerve centre that enables relatively very complexly coordinated movements to be performed, such as a starfish's movements to open the shells of bivalves. Along two other major lines of evolution (from the primitive worms to crustraceans and spiders, and from primitive worms to insects) a more massive anterior... ganglion becomes formed which subordinates the working of lower-lying ganglia to itself (see Figs. 15-17).
The origin of this type of nervous system is conditioned by differentiation of a leading organ along with other sense organs, which thus becomes the main organ mediating the organism's vital activity. [p. 163]
The evolution of such a ganglial nervous system took the direction of increasing differentiation, which was associated with segmentation of the animal's body.
 The change of activity within this stage of evolution consisted in its ever greater complication, which occurred along with the evolution of animal's organs of [sense] and action and nervous system. Both the general type of structure of activity, however, and the general type of reflection of the environment did not alter greatly throughout this stage of evolution. Activity is stimulated and controlled by the reflection of a number of separate properties; [their reciprocal sensitivity toward] reality is consequently never perception [p. 164] of the wholeness of things. In the least organised animals (e.g., worms) activity is always stimulated by the effect of one property of some sort, so that, for example, a characteristic feature of their search for food is that it is always made (as Wagner [(1928)] says) 'by means of some one sense organ, without the assistance of other sense organs: touch, more rarely smell or vision, but always only one of them'.
The change of activity within this stage of evolution consisted in its ever greater complication, which occurred along with the evolution of animal's organs of [sense] and action and nervous system. Both the general type of structure of activity, however, and the general type of reflection of the environment did not alter greatly throughout this stage of evolution. Activity is stimulated and controlled by the reflection of a number of separate properties; [their reciprocal sensitivity toward] reality is consequently never perception [p. 164] of the wholeness of things. In the least organised animals (e.g., worms) activity is always stimulated by the effect of one property of some sort, so that, for example, a characteristic feature of their search for food is that it is always made (as Wagner [(1928)] says) 'by means of some one sense organ, without the assistance of other sense organs: touch, more rarely smell or vision, but always only one of them'.
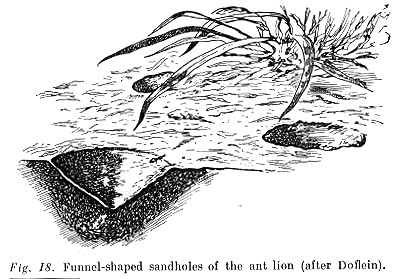
The complicating of activity within the limits of this common type takes two main directions, one of which is most clearly expressed in the line of evolution leading from worms to insects and spiders. It is manifested in the activity of animals acquiring the character of sometimes very long chains consisting of a great number of reactions corresponding to separate consecutive influence. A vivid example is the often cited behaviour of the larva known as the ant lion.
The ant lion buries itself in sand just so deep that grains of sand begin to touch the surface of its head, which causes a jerking backthrusting of the head and anterior part of the body that throws the sand upward. As a result a funnel of regular shape is formed in the sand at the centre of which the ant lion's head juts out. When an ant gets into the funnel it [p. 165] inevitably causes a few grains of sand to slide down. These, falling on the ant lion's head, trigger off the 'throwing' reflex described. Some of the sand thrown up hits the ant, which slides down to the bottom of the funnel with the falling sand. Then, as soon as the ant touches the jaws of the ant lion, they shut on it and the victim is sucked dry (See Fig. 18, after Doflein, simplified).
The mechanism of this activity is one of elementary [sensory-motor] reflexes -innate unconditioned and conditioned.
 Activity of this kind is particularly characteristic of insects in which it attains its highest degree of development. This line of the complicating of activity is not progressive and does not lead to its subsequent qualitative alteration.
Activity of this kind is particularly characteristic of insects in which it attains its highest degree of development. This line of the complicating of activity is not progressive and does not lead to its subsequent qualitative alteration.
The other line followed by the complicating of activity and sensitivity, on the contrary, is progressive. It leads to a change in the structure of the activity itself and on that basis to the rise of a new form of reflection of the environment characterised by an already higher, second stage in the evolution of animal's psyche, namely that of the perceptive psyche. This... trend is linked with the progressive line of biological evolution (from wormlike animals to primitive chordates and then to vertebrates).
The [transitional] complicating of animal's activity and sensitivity is expressed here in their behaviour being governed by a combination of many simultaneous influences. Examples can be drawn from the behaviour of fish. A marked contradiction is distinctly observable in them between an already relatively very complex content of processes of activity and a high development of separate functions on the one hand, and a still primitive structure of this activity on the other hand.
Let us turn again to special experiments.
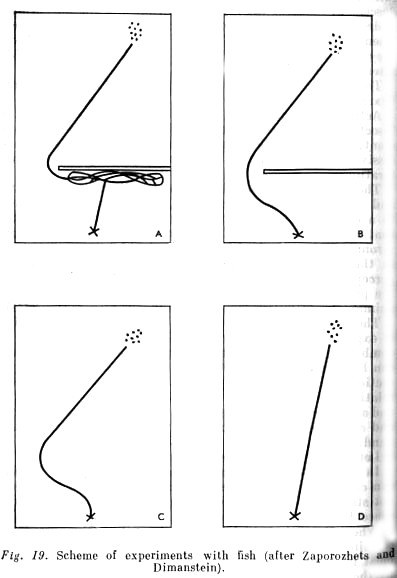
In an isolated aquarium in which live two young American catfish, a white cheesecloth partition is fixed that does not stretch as far as one of the walls, so that there is a free space between it and the wall.
When the fish, which usually keep together, are in a certain end of the aquarium (always the same one), a bit of meat is dropped to the bottom of the other end. Attracted by the spreading flavour of meat the fish make directly for it along the very bottom. In doing so they come up against the gauze partition; having approached to within a few [p. 166] [p. 167] millimetres of it, they remain for a moment seeming to regard it, and then swim along it from one side to the other, until by chance they find themselves opposite that gap at the side, through which they can penetrate into the part of the aquarium where the meat is.
The fish's observed behaviour is due to two main influences. It is excited by the scent of meat and develops in the direction of this main, dominant influence. On the other hand the fish [see] (visually) a barrier, so that their movement toward the spreading scent takes on a complicated, zigzag character (see Fig. 19 A). This is not, however, a simple chain of movements: at first there is a reaction to the stretched gauze and then a reaction to the scent. There is also no simple merging of these two influences giving rise to a resultant movement. It is complexly coordinated activity in which a twofold content can be objectively distinguished: (1) a definite direction of [approach] activity leading to an appropriate result (this content arises under the influence of the scent, which has the biological sense of food for the animal); (2) turning movements proper (the content of which is associated with a definite influence -the barrier- but this influence is distinguished from the effect of the smell of food, and cannot independently excite the animal's activity; the gauze in itself does not cause any reaction in the fish). This second effect is not associated with an object that excites activity and toward which it is directed, but with the conditions in which this object is presented. This is an objective difference between the two effects and is their objective relationship. Is the objective relationship, however, reflected in the activity of the fish being studied?.... Does the one operate as associated with the object, i.e. with what stimulates activity? And the second as relating to the conditions of the activity, in general as an other?
To answer that let us continue with the experiment.
As the experiments in feeding the fish in the presence of a barrier on their path to the food continued, there is a kind of gradual 'melting away' of surplus movements so that finally the fish immediately move toward the gap between the gauze and the wall of the tank, and then to the food (Fig. 19, B).
Let us now pass to the second part of the experiment. In it we remove the barrier before feeding the fish. Although [p. 168] it was located quite close to the initial point of the fish's movement, so that they could not help noticing its absence in spite of their relatively weak vision, nevertheless the fish fully followed the roundabout path, i.e. moved as they would if the barrier had been in place (Fig. 19 C). Subsequently their path straightened, of course, as is shown by Fig. 19, E, but that happened only gradually.
Thus the effect governing the roundabout movement was firmly associated in the studied fish with the effect of the food itself, with its smell. That means that it [the barrier] was [sensed] by the fish... not as... another thing [but “from the start continuously with the smell of food”].
Thus, as the result of a gradual complicating of... [motor] activity and sensitivity, we observe the rise of a... [preadaptive] nonconformity, a [objective] contradiction in their behaviour.... In other words... [the observable] activity [of the “fish (and seemingly of certain other vertebrates)"] is governed in fact by an influence already coming from separate things (food, a barrier), while the reflection of reality remains a reflection in them of the [sensory] aggregate of its different properties.
During subsequent evolution this nonconformity [between sensory reflection and preadaptive leading activity] was resolved through a change in the [predominant] form of reflection and a further restructuring of the animal's general type of activity; there was a transition to a new, higher [(qualitatively different)] stage of development of reflection [i.e., the “perceptual stage” of psyche].
Before we start to examine this new stage, however, we must first dwell on yet another special problem that arises in connection with the general issue of the variability of animal's activity and sensitivity.
This is the matter of so-called instincts, i.e. of innate, unconditioned reflex behaviour and of behaviour that is altered by the effect of an animal's external conditions of existence, and by its individual experience.
Views that link the successive stages in the evolution of the psyche with these different mechanisms of animal's adaptation to their environment are very common in psychology. The lowest stage in the evolution of the psyche is pictured, from this point of view, for instance, as behaviour [p. 169] based on animal's so-called tropisms or instincts; higher stages of evolution are formed by individually alterable behaviour, i.e. behaviour built on conditioned reflexes.
These views are based on the indisputable fact that the higher we go up the ladder of biological evolution, the more and more perfected is... adaptation to variability of the environment, the more dynamic their activity becomes, and the easier it is for animals to 'learn'. The concrete understanding of the development of animal's behaviour, however, advanced by the holders of this point of view, is extremely simplified and is, in essence, incorrect.
First, of all, there is no foundation for counterposing [i.e., to place in opposition], as different genetic stages of behaviour, (a) inherited behaviour, allegedly unalterable by external influences, and (b) behaviour built up in the course of an animal's individual development, in the course of its individual adaptation. 'Individual adaptation', Pavlov [(1949)] said, 'exists throughout the animal world.'
The counterposing of innate and individually adapted behaviour came about, on the one hand, from incorrect reduction of the mechanisms of animal's behaviour to its innate mechanisms, and on the other hand from the old idealist understanding of the term 'instinct'.
Tropism is usually considered the simplest form of innate behaviour. The theory of tropism in relation to animals was developed by Loeb. According to him [(1918)] a tropism is a forced, automatic movement caused by a dissimilarity of physico-chemical processes in the symmetrical parts of an organism as a consequence of the one-sidedness of influences affecting it.
 An example of this forced and invariable movement is the growth of the roots of a plant, which is always downward no matter in what position we place the plant. Similar phenomena can also be observed in animals; it does not follow from that, however, that the activity of these animals boils down to a mechanism of tropisms and that it is not plastic, altering through the effect of experience.
An example of this forced and invariable movement is the growth of the roots of a plant, which is always downward no matter in what position we place the plant. Similar phenomena can also be observed in animals; it does not follow from that, however, that the activity of these animals boils down to a mechanism of tropisms and that it is not plastic, altering through the effect of experience.
We know, for example, that most Daphnia possess positive [p. 170] phototropism, i.e. that they perform forced movements toward light. The behaviour of Daphnia, however, as the special experiments of Blees [(1919)] and of Soviet workers (Leontyev and Basin) have shown, in no way resembles the 'behaviour' of plant roots.
Blees's experiments were made as follows.
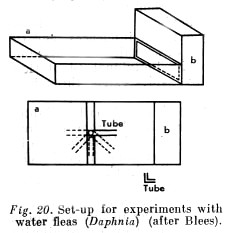
A small, flat aquarium, illuminated from one side only, was used. A glass tube, bent at a right angle, was fixed in the centre of the tank so that one end lay horizontal under the water and the other end projected vertically out of the water (Fig. 20).
 At the start of the experiments the horizontal limb pointed toward the illuminated wall of the tank, i.e. toward the light source (positioned as shown in Fig. 20).
At the start of the experiments the horizontal limb pointed toward the illuminated wall of the tank, i.e. toward the light source (positioned as shown in Fig. 20).
Water fleas (Daphnia) were pipetted and put into the tube; they quickly passed down the vertical part of the bend and began at once to move along the horizontal limb toward the light. On emerging from the tube they then swam freely to the illuminated wall of the tank. Their behaviour thus remained strictly controlled by the effect of light.
In the next experiments the tube was rotated by 45 degrees from the direction of the light (as shown by the dotted lines in Fig. 20).
In these conditions the water fleas came out of the tube as before but more slowly.
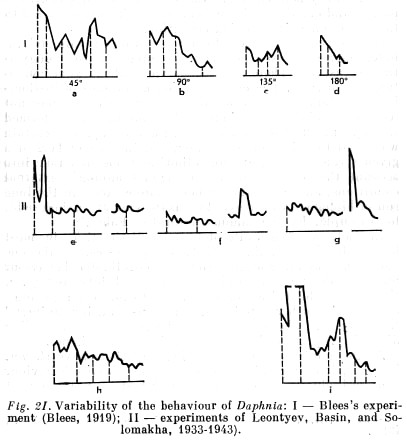
That fact is also easily explainable from the angle of tropism. It can be supposed that we have a summation of two directions here, the influence of the light and the influence [p. 171] of the wall of the tube, obstructing direct movement, and now pointing a bit to one side. The summation of these two directions is also expressed in the fleas' slow passage through the tube. Repetition of [exposure to this condition]..., however, demonstrated that the fleas' passage through the tube was quicker and quicker [across trials], until finally it was close to the speed needed to negotiate the tube when it pointed directly to the light. A certain training was consequently observed in Daphnia, i.e. their behaviour was gradually adapted to the given conditions.
In the next experiments the tube was turned to 90 degrees, then to 135 degrees, and finally to 180 degrees. In all these positions the water fleas also gradually learned to pass quite quickly out [p. 172] of it, although they were forced in the last two cases to move away from the light in a direction opposite to their tropism (Fig. 21).
That fact can also be shown at first glance not to contradict the 'forced character' of phototropism in Daphnia; it can be supposed that their positive tropism is converted into negative tropism by an influence unknown to us. That supposition, however, is refuted by the fact that after emerging from the tube the fleas again moved toward the light.
Thus, as follows from the facts adduced, the behaviour of water fleas by no means consists in machinelike, forced movements or tropisms. The tropisms of animals are not elements of a wholly mechanical behaviour, but mechanisms of elementary behaviour processes, behaviour that is always plastic and capable of being reorganised in accordance with the environment's changing conditions.
Another concept with which the notion of animal's innate, rigorously fixed behaviour is linked in psychology is that of instinct. There are various views about what constitutes an instinct. The most common is the idea of instinctive behaviour as behaviour that is inherited and does not require learning of any kind, behaviour that is performed through the effect of certain stimuli and always in a certain way, and that is completely identical in all members of a given species. It is therefore 'blind' and does not take into account the features of the individual animal's external conditions, and is capable of being altered only in the long run of biological evolution. That is the view of instinct held for example, by the famous naturalist Fabre [(1910)].
In fact, we can quite definitely distinguish, in most highly developed animals, between processes, on the one hand, such as are manifestations of consolidated behaviour built up during the species' history and inherited (e.g. the innate 'ability' of certain insects to build honeycomb), and on the other hand, such as arise during animal's 'learning' (e.g. bees learning to choose correctly a feeding dish of syrup marked by a figure of a certain shape).... [p. 173]
.... Animal's behaviour is... species behaviour, but it is at the same time very plastic.
Rigorously fixed instinctive behaviour is thus by no means the initial stage in the development of animal's behaviour. That is the first point.
The second point is that there is also no instinctive behaviour in... activity at higher stages of evolution such as is unalterable by the effect of an animal's individual living conditions. That means, strictly speaking, that there is no behaviour in general that is fixed once and for all, and that follows only a ready-made pattern laid down in advance.... [p. 174]....
Even more obvious is the fact that animal's so-called individual behaviour is shaped in turn on the basis of species instinctive behaviour and cannot otherwise arise.... We can only affirm that innate mechanisms play a greater role in some animals and mechanisms of individual experience in others. This [merely quantitative] difference, however, also does not reflect the real... [i.e., essential, proper, generally useful, or differentiating] character of the evolution of the psyche in the animal world. Rather it indicates a peculiar... characteristic of [some rather limited] lines of animal evolution. Innate behaviour is most clearly manifested... in insects, which are known to be located on a side branch of evolution.
A difference in the type of mechanisms that implement animal's adaptation to changes in the environment [e.g., the relative preponderance of reflex, instinct, or reason] thus [p. 175] cannot serve as the sole criterion of the evolution of their psyche. It is not only in what main way... behaviour is altered that is important but [more] primarily what its content itself and inner structure are and what are the forms of reflecting reality that are naturally associated with them.
2. The Stage of the Perceptive Psyche
The next stage after that of the elementary sensory psyche, the second stage of evolution, can be called that of the perceptive psyche. It has the capacity to reflect external, objective reality already in the form of a reflection of things rather than in the form of... sensations evoked by separate... or a combination of properties.
The transition to this stage in the evolution of the psyche is associated with a change in the structure of animal's activity already prepared for in the preceding stage. This change consists in the content of this activity already mentioned above, which is [in the sensory psyche]... related to the conditions in [i.e., properties of]... the environment, rather than to the object itself toward which the animal's activity is directed....
When a mammal is separated from food by an obstacle, it will, of course, go around it.... Between the activity of fish [in the obstruction tank] described [above] and that of mammals, however, there is a great difference, which is expressed in this, that while the content of the fish's activity (roundabout movements) was retained after removal of the barrier and disappeared only gradually, higher animals usually make directly for the food in such a case. This means that the influence to which mammal's activity is directed no longer merges with influence from the barrier in them, but both operate separately from one another for them. The direction and end result of the activity depends [p. 176] on the former, while the way it is done, i.e. the mode in which it is performed (e.g. going around the obstacle) depends on the latter. This special [modal] make-up or aspect of activity, which corresponds to the conditions in which the object exciting it is presented, we shall call operation.
It is this [selective and observable] distinguishing of [appropriate] operations in activity that indicates that properties affecting an animal... [are beginning] to fall into groups: on the one hand... properties... that characterise the object to which the activity is directed, while on the other hand properties... that determine the mode of the... operation [performed]. Whereas differentiation of the affecting properties was linked at the stage of the elementary sensory psyche with their... uniting around the dominant stimulus, the integrating of the affective properties into a single integral image, and their unification as the properties of one and the same thing now arise for the first time. The surrounding reality is now reflected by the animal in the form of more or less separated images of separate things.
The majority of now existing vertebrates are at various levels of that stage of the perceptive psyche. The transition to this stage was seemingly linked with the passage of vertebrates to a terrestrial mode of life.
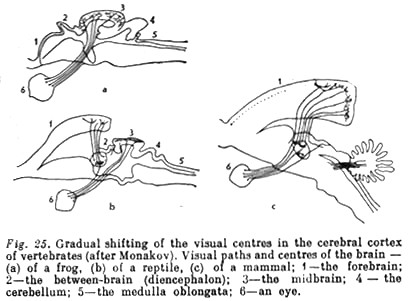
The rise and development of a perceptive psyche... were governed by several essential anatomical and physiological changes. The main one was the development and change of role of distant sense organs (i.e. ones operating at a distance), primarily of vision. Their development was expressed in an alteration both of their significance in the general system of activity and in the form of their anatomical interconnections with the central nervous apparatus. Whereas differentiation of sense organs in the preceding stage of evolution had led to the singling out of dominant organs among them, among vertebrates the leading organs more and more became those that integrate external influences. That became possible because of the simultaneously occurring restructuring of the central nervous system and the formation of a forebrain, and then of a cerebral cortex (for the first time in reptiles). Originally (among fish, amphibians, and reptiles) the forebrain was a purely olfactory formation constituting a sort of continuation of their central olfactory apparatus. In subsequent evolution (among [p. 177] [p. 178] mammals) the importance of the olfactory centres in the cerebral cortex was greatly reduced through the representation of other sense organs. This is clearly seen when we compare the place occupied by the olfactory cortex... in a hedgehog (Fig. 22) and a monkey (Fig. 23).
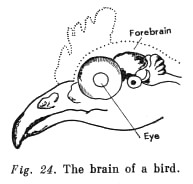 Vision, on the contrary, whose 'corticalisation' occurred initially with the reptiles, occupies a relatively ever greater place in the cortex (see Fig. 25). In birds the eyes become the main receptor (Fig. 24). Vision also plays the main role in many higher mammals.
Vision, on the contrary, whose 'corticalisation' occurred initially with the reptiles, occupies a relatively ever greater place in the cortex (see Fig. 25). In birds the eyes become the main receptor (Fig. 24). Vision also plays the main role in many higher mammals.
The organs of external movement developed simultaneously, i.e. animal's 'natural tools' enabling them to perform the complicated operations demanded by life in a terrestrial environment (running, climbing, pursuing prey, overcoming obstacles, etc.). Animal's motor functions were also [p. 179] more and more corticalised (i.e. transferred to the cortex of the brain), so that full development of operations proceeded in animals in connection with the evolution of the cortex.
Thus, whereas the activity of lower vertebrates was still mainly linked with lower-lying centres (subcortical ganglia), it subsequently became more and more dependent on the cortex, changes in whose structure also reflect all its subsequent evolution.
 Differentiation of the operations that characterise the stage of the perceptive psyche laid the basis for the evolution of a new form of fixing animal's experience, for fixing it in the form of motor habits [i.e., automatic motor operations] in the narrow sense of the term.
Differentiation of the operations that characterise the stage of the perceptive psyche laid the basis for the evolution of a new form of fixing animal's experience, for fixing it in the form of motor habits [i.e., automatic motor operations] in the narrow sense of the term.
Any connections arising in individual experience [whether automatic or intentional] are sometimes called habit [e.g., see William James on Habit as “the enormous flywheel of society", Vol. 1, p. 121)]. In the wide conception, however, the concept of habit becomes blurred, embracing a vast circle of quite different processes, beginning with changes in the reactions of infusoria and ending with man's complicated actions. In contrast to that, in no way justified broadening of the concept of habit, we shall call only fixed [i.e., acquired and then automated] operations habits.
That definition of habit coincides with the understanding... first advanced in Soviet psychology by Protopopov [(1935)], who demonstrated experimentally that motor habits are formed in animals from the motor elements of overcoming obstacles, and that the content of habits is determined by the character of the obstacle itself, while the stimulus (i.e. main exciting influence) only affects a habit dynamically (the speed and firmness of its forming) and is not reflected in its [modal] content.
The motor elements forming part of the habits of animals may be different in character; they may be both species, innate movements and movements acquired in previous experience; finally they may be movements fixed [i.e., in sequence or structure] in the course of the chance motor trial and error made by the animal during the formation of the given habit.
Clearly expressed habits in the [higher] sense [i.e., those acquired by way of initial effort or practice and only then automated into 'habitual' operations] are observed at first only in animals that have a cerebral cortex. The mechanism of the formation and fixing of systems of precisely cortical conditioned nerve connections must therefore [p. 180] be considered the physiological base of the formation of [such adaptive and preadaptive] habits.
[What Leontyev is hinting at here is that the cortex initially arose and continues to function not only adaptively (i.e., in the role of allowing the organism to be more attentive to important environmental contingencies) but is also preadaptive of further psychical evolution because it provides a physiological means by which learned perceptually attentive or motor “behaviours” -i.e., actions with respect to biologically significant “objects” or “obstacles"- become automated into “operations” controlled by the basal ganglia. This “habitual” automation thereby frees the cortex to attend to other and newer significant contingencies (whether they be “represented” in immediate perceptual experience, remembered, or anticipated).]
The problem of the existence of [reflective] representations in animals is still a matter of dispute. A vast number of facts, however, convincingly indicate that animals [at the perceptual level of psyche and beyond] have representations.
Tinklepaugh... [(1928)] ... showed fruit to an animal (monkey) and then, behind a board, surreptitiously replaced it [with] lettuce, which is much less attractive. The animal was then allowed to move behind the partition; though finding lettuce there it nevertheless continued to look for the fruit...
Similar experiments made with a fox by Voitonis and Kreknina [(1935)] yielded the same results.
The observations on a dog described by Beritov [(1934)] are of great interest in this respect. In his experiments with conditioned reflexes, the dog was put in a certain [starting place] and then given a conditioned signal in response to which it ran to a simultaneously uncovered feeding dish, and received food. During these experiments the following test was made: before the dog was brought into the laboratory, it was walked to the far end of the corridor and shown food lying there, without however being signaled to take it. Then it was led back to the laboratory and given the conditioned signal. When it ran to the feeding dish, however, there was no food there. ...[T]he dog did not return as usual to its [starting] place but ran out into the corridor to the spot where it had previously seen food.
Buytendijk and Fischel's [(1930)] experiments with dogs were of a more specialised character. They were able to demonstrate [p. 181]... that in contrast to lower vertebrate organisms (fish), a dog is [able to remain persistently] oriented [toward a] previously experienced... [rewarding lure]... ([now]... concealed from... sight) [during its new] reactions to [experimentally presented objects]...
Thus, together with a change in the structure of animal's activity and a corresponding change in the form of their reflection of reality there is also a restructuring of the function of memory. Earlier, at the stage of elementary sensory psyche, this [retentive] function was expressed in the motor sphere... under the impact of external influences, ...and in the sensory sphere in... the links between separate effects. Now, at this higher [perceptive psyche] stage [and beyond]..., the mnemonic function operates in the motor sphere in the form of motor habits, and in the sensory sphere in the form of a primitive, image memory.... [p. 182]....
The main change in the processes of differentiation and generalisation during the transition to a perceptive psyche is expressed in the rise in animals of [the new reflective capacity to utilize]... the images of [biologically significant] things [i.e., as unified though distinguishable entities]. [Organisms at the stage of perceptive psyche can differentiate between objects (as things) and reflect upon their respective biological significance. It is by way of this new perceptual means that these organism's ability to learn (i.e., to make generalizations and to adapt those generalizations according to new environmental circumstances) is broadened in scope.]
The origin and evolution of [such] generalised reflection of things is already a much more complex problem, on which we must dwell specially.
The [perceptual] image of a thing is not a simple sum of individual sensations or the mechanical product of many simultaneously operating properties belonging to objectively different things. If, for instance, we have two [objects] of whatever sort A and B that possess properties a, b, c, d and m, n, o, p, then for an image to arise, these... properties must function as part of two separate entities (A and B), i.e. they must be differentiated in precisely that respect. This means, also, that when the given influences are repeated among others, their previously differentiated unity must be perceptible as the thing itself. Given the inevitable variability of the environment, however, and of the conditions of perception itself this is only possible when the image of the thing arising is generalised.
In the cases described we see dual interconnected processes: those of the transfer of operations from one concrete situation to another, objectively similar to it, and those of the forming of a generalised image of a thing. The generalised image of the thing, in arising together with the shaping of an operation in relation to, and on the basis [p. 183] of, this thing, enables the operation to be transferred subsequently to a new situation; in this process the previous operation comes into a certain disharmony with the... conditions of [the new] activity... and... is altered and reorganised. The generalised image of the thing is correspondingly reorganised, made more precise, and absorbs the new content as it were, which in turn leads to the possibility of a further transfer of the [mastered] operation to new... conditions [again] calling for... even fuller... reflection by the animal.
 Perception is thus still fully included... in the animal's external motor operations. Generalisation and differentiation, synthesis and analysis take place in a single process.
Perception is thus still fully included... in the animal's external motor operations. Generalisation and differentiation, synthesis and analysis take place in a single process.
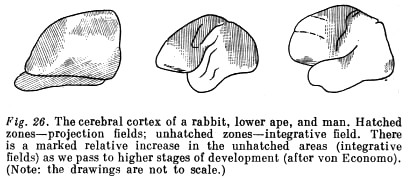
The evolution of operations and generalised perception... finds its reflection in a further complicating of the cerebral cortex. There is further differentiation of the integrative fields, which occupy an ever bigger place in the cortex (see Fig. 26).
The function of these higher integrative fields is, ... to integrate separate influences.
3. The Stage of [Animal] Intellect
The psyche of most mammals remains at the stage of the perceptive psyche, but the most highly organised mammals have risen to an even higher stage of [mental] evolution. [p. 184] [Organisms at the stage of Animal intellect can reflect upon objects and relations (including social relations) between other organisms. It is by way of this new intellectual means that their ability to learn these relations (i.e., to make generalizations and to also adapt those generalizations to new circumstances) is broadened in scope.]
This new, higher stage is normally called the stage of intellect (or 'manual thinking').
The intellect of animals, of course, is not quite the same as human reason; as we shall see, there is an immense qualitative difference between them.
The stage of [animal] intellect is characterised by very complex... forms of reflecting reality. Therefore, before we deal with the conditions for the passage to this stage, we must describe the activity of animals that are at this stage... in its external expression.
The [externally observable] intellectual behaviour of the most highly developed animals -the anthropoid apes- was first systematically studied in the experiments carried out by Köhler [(1925)]....
The apes (chimpanzees) were housed in a cage. Outside the cage, just far enough away that the ape's arm could not reach it, bait was placed (bananas, oranges, etc.). Inside the cage there was a stick. The ape, attracted by the bait, could only bring it closer to itself in one way, by using the stick. How did the ape behave in this situation? As it happened, it first began to try and snatch the bait directly with its hand. The attempts were unsuccessful. The ape's activity seemed to fade for a time. It turned away from the bait and stopped its attempts. Then activity was resumed, but now took another path. Without trying to grab the fruit directly by its hand, the ape picked up the stick, thrust it toward the fruit, touched it, drew the stick back, again thrust it out and again drew it back, with the result that the fruit was drawn closer and the ape snatched it up. The problem was solved.
The many other problems set anthropoid apes have been built on the same principle; their solution also required the adoption of a mode of activity such as could not be formed during solution of the problem set. For example, bananas were hung from the upper lattice of the enclosure where the apes were kept, out of their direct reach. Nearby was an empty box. The only possible way of reaching the bananas in this case was to drag the box over to the spot above which the bananas hung, and to use it as a stand. Observations showed that apes solved this problem without noticeable preliminary learning.
Thus, while operations are formed slowly at a lower stage of evolution, by way of many trials during which successful [p. 185] movements are gradually fixed [i.e., established, mastered], and other, unnecessary movements are gradually inhibited, and fade out, in the case of apes we observe first a period of complete failure -many attempts not leading to accomplishment of the activity, and then suddenly, as it were, the finding of an operation that almost immediately leads to success. That is the first characteristic feature of the intellectual activity of [such] animals.
A second characteristic feature is that when an experiment is repeated once more, the [previously successful] operation concerned is reproduced [i.e., selected], in spite of its having been performed only once, i.e. the ape solves a similar problem... without any preliminary trials.
 A third feature of this activity is that ...[a given] solution... is... [selectively] transferred by the ape to... conditions only [suitably] similar to those in which [it] was first found. If an ape, for example, is now deprived of the stick, it easily employs some other suitable object in place of it. If the position of the fruit is altered in relation to the cage, or if the situation is altered slightly... the animal all the same finds the necessary solution. The solution, i.e. the [appropriate] operation, is [selected,] transferred... and adapted to this new situation...
A third feature of this activity is that ...[a given] solution... is... [selectively] transferred by the ape to... conditions only [suitably] similar to those in which [it] was first found. If an ape, for example, is now deprived of the stick, it easily employs some other suitable object in place of it. If the position of the fruit is altered in relation to the cage, or if the situation is altered slightly... the animal all the same finds the necessary solution. The solution, i.e. the [appropriate] operation, is [selected,] transferred... and adapted to this new situation...
We must note.... that anthropoid apes are capable of uniting two different operations into a single [problem solving action].
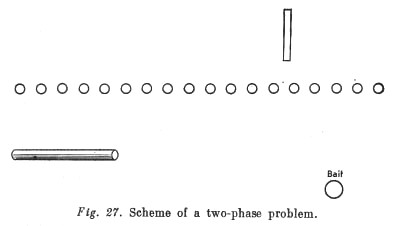
For example, bait is placed outside the cage in which the animal is housed, at a certain distance from it. Rather nearer to the cage but also beyond the animal's reach is a long stick. Another shorter stick that can reach the long one but not the bait is put into the cage. To solve this problem the ape must first pick up the short stick, draw the long stick to itself, and then pull the bait to it with the long stick (see Fig. 27). Apes usually cope with such 'two-phase' tasks without special difficulty. So a fourth feature of [their] intellectual activity consists in a capacity to solve two-phase tasks.
Subsequent experiments by other researchers have shown that these characteristic features are preserved as well in [p. 186] the more complicated behaviour of anthropoid apes (Ladygina-Kots and Vatsuro [, 1928]).
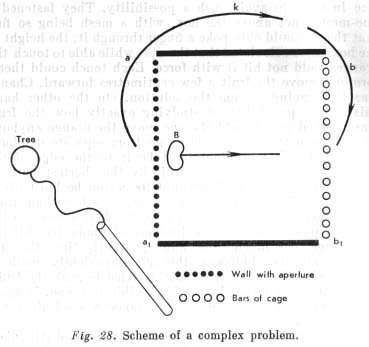 An example of the solution of a very complicated task by an anthropoid ape is the following experiment (Fig. 28). In the enclosure where the apes lived a box was set one side of which was a [barred] cage [(b1)] while the other side had a narrow, longitudinal slit [(a1)]. Fruit [(B)] was put near the back of this box, clearly visible both through the bars at the front, and thorough the slit behind. The bait was too far away from the bars for the ape to reach it. It was also impossible to reach the bait from the rear wall itself because the slit [(aperture)] was too narrow to admit the ape's arm. A strong stake [(Tree)] was driven into the ground near the rear wall, and a stick fastened to it by a not very long chain.
An example of the solution of a very complicated task by an anthropoid ape is the following experiment (Fig. 28). In the enclosure where the apes lived a box was set one side of which was a [barred] cage [(b1)] while the other side had a narrow, longitudinal slit [(a1)]. Fruit [(B)] was put near the back of this box, clearly visible both through the bars at the front, and thorough the slit behind. The bait was too far away from the bars for the ape to reach it. It was also impossible to reach the bait from the rear wall itself because the slit [(aperture)] was too narrow to admit the ape's arm. A strong stake [(Tree)] was driven into the ground near the rear wall, and a stick fastened to it by a not very long chain.
The solution of this problem consisted in pushing the stick through the slit in the rear wall and shoving the fruit forward to the front bars, through which it could then be pulled out simply by the hand.
How did the ape behave in this situation? On coming up to the cage and noticing the fruit, it first tried to reach it thorough the bars. Then it went round the box and looked at the fruit thorough the slit at the back. It tried to pull the fruit through the slit by means of the stick, but that [p. 187] was impossible. Finally, the animal pushed the fruit away with the stick, and went round the box so as to pull it out through the bars.
How were all these complicated operations, observed in the experiments described, formed? Did they really originate suddenly without any preliminary preparation, as it seemed from the first outward impression? Or were they built up in the main in the same way as in the preceding stage of evolution, i.e. through gradual, albeit much faster, selection and fixing of movements leading to success?
The answer to that is clear from an experiment describe by French workers, which was carried out as follows. An anthropoid ape was housed in a cage. A small box was fixed on the outside of the bars with an opening on the side opposite the bars. An orange was put close to the wall of the box. To get it in this situation the animal had to knock it out of the box by a blow. Since such a blow could be made accidentally, the researchers adopted the following clever device [p. 188] in order to avoid such a possibility. They fastened a fine-meshed net above the box, with a mesh being so fine that the ape could only poke a finger through it; the height of the box was so calculated that the ape, while able to touch the orange, could not hit it with force. Each touch could therefore only move the fruit a few centimetres forward. Chance was thus excluded from the solution. On the other hand this gave a possibility of studying exactly how the fruit was knocked out. Would the ape move the orange anyhow, so that its path would accidentally take it to the edge of the box? Or would it guide the fruit by the shortest path to the exit from the box, i.e. would its action be built up of movements directed in a definite way rather than from chance ones? The animal itself gave the best answer to the question posed. Because the business of gradually shifting the orange took much time, and apparently tired the animal, it already, halfway through impatiently made a searching movement of the arm, i.e., tried to grab the fruit; having discovered that it was impossible to do so, it again began slowly pushing it until the orange was within range of its hand [(Guillaume & Meyerson, 1930)].
Köhler [(1925/57)] considered that the main attribute distinguishing the behaviour of these animals from that of other members of the animal kingdom, and which brought it closest to the behaviour of man, was precisely that their operations were not shaped gradually through trial and error but arose suddenly, independently of previous experience, by insight as it were. A second attribute of intellectual behaviour, derived from the first, he considered to be a capacity to remember the found solution 'once and for all', and to transfer it broadly to other conditions similar to the original ones. As regard the fact of ape's solving two-phase problems, Köhler and other following him consider that a combination of two moments underlies it: the animal's 'insight' and the transfer of a solution earlier found. They thus did not consider [the two-phasedness of the experimental tasks set for the apes -i.e. the necessity of calling upon previously established operations in order to perform a novel, larger, problem-solving action] to have any fundamental [theoretical-explanatory] significance. [p. 189]
From [their] point of view, it is sufficient to explain the main fact, i.e. the fact of an animal's sudden finding of a way to solve the first, initial problem, in order to understand the whole peculiarity of ape's intellectual activity.
Köhler tried to explain [the fact of insight] by [appeal] to the apes having a faculty of correlating separate things, distinguishable from one another, in perception, so that they were perceived as part of a single 'integral situation' (Gestalt).
This property of perception itself, its structured character, is only a partial case, in Köhler's view, expressing the general 'Gestalt principle' that allegedly underlies not only the psyche of animals and man and their vital activity, but also the whole physical world.
From that point of view the 'Gestalt principle' can serve as an explanatory principle, but itself is then inexplicable and does not require explanation. The attempt to bring out the essence of intellect starting from this idealist 'Gestalt theory' is, it goes without saying, unsound. Quite clearly it is not sufficient to enlist the structured character of perception to explain the peculiarity of higher animal's behaviour. For, from the standpoint of the adherents of the 'Gestalt principle', structured perception is not only peculiar to the higher apes but is also peculiar to much less developed animals; intelligent behaviour, however, is not observed in the latter. [Stated plainly, the failure of the Gestalt account to recognize the importance of two-phase actions blurred the observable dividing line between perceptive psyche and higher animal intellect.]
This [Gestalt form of] explanation is also unsatisfactory from another aspect. By stressing the suddenness of the intellectual solution and isolating that fact from the content of an animal's [prior] experience, Köhler left a whole number of circumstances out of account that characterise the behaviour of apes in their natural environment. [Furthermore, in its isolated emphasis on the suddenness of the ape's problem-solving skills, a false dividing line is laid down by the Gestalt theory.]
Buhler [(1930)] it seems was the first to draw attention to the fact that there is something in common between an ape's drawing a fruit to itself by means of a stick, and pulling a fruit growing on a tree to itself by means of a branch. Attention was then drawn to the fact that the roundabout path observed in apes could also be explained by... these animals, living in forests and passing from one tree to another, must constantly 'orient themselves' to the route in advance, or else they would find themselves in an impasse of the natural labyrinth formed by the trees. It is not accidental, therefore, that apes display a developed [p. 190] faculty for solving problems in a 'roundabout way'.
The idea that the explanation of ape's intellectual behaviour must be sought above all in its link with their normal species behaviour in their natural environment has been expressed more and more definitely of late in the works of psychologists and physiologists.
From that [additive, reductionist,] point of view [, however,] an intellectual 'solution' is nothing more than the application in new conditions of a mode of activity phylogenetically developed. [According to their theoretical portrayal of the data collected in animal experiments]... this transfer of a mode of action differs from the ordinary transfer of operations in other animals only in happening within [quantitatively] wider limits.
Thus, according to this conception of the intellectual behaviour of apes [e.g., Skinner's "operant” behaviorism],.... the sudden solution of an experimental problem must itself be understood as the result of the animal's capacity for a broad transfer of operations.
That [additive] conception... has the virtue that it does not counterpose the animal's intellect to either its individual or species experience, and does not separate intellect from habit. But it also comes up against serious difficulties. First of all it is clear that neither the moulding of operations nor their transfer to new conditions of activity can serve as distinguishing attributes of the behaviour of higher apes, because both... are common to animals at a lower stage of evolution. We observe both.... in many other animals as well, viz., among mammals and birds. [This aspect of the] difference in activity and psyche between the latter and apes, it turns out, is a purely quantitative one: a slower or quicker moulding of the operation, and narrower or broader transfers. But the behaviour of apes differs qualitatively as well from that of lower mammals. Their use of instruments and the special character of their operations are quite clear evidence of that. [p. 191]
Furthermore, the conception of animal's intellect cited above leaves the main thing undisclosed, namely what is the wide transfer of actions observed in apes and what is the explanation of it.
To answer these questions we must... make a... characteristic fact that in [Köhler's] opinion is of no fundamental importance the starting point of [our] analysis, namely ape's capacity to solve two-phase problems.
In [the first phase of] two-phase problems.... it is necessary... to push the fruit away.... Touching the stick by itself leads to taking hold of it but not to seizure of the fruit that attracts the animal.... Unconnected with the next phase it lacks any biological sense whatsoever. It is a phase of preparation. The second phase -use of the stick- is already the phase of the realisation of [action] toward a goal, directed to satisfying a given biological need of the animal. Thus, if we approach the ape's solution of any of the problems given them by Köhler from this point of view it proves that each of them required two-phase [actions]: to pick up a stick-to pull the fruit to itself, to move away from the bait-to possess the bait, to turn the box over-to reach the fruit, and so on.
What is the essence of these two phases of the ape's [actions]? The first, preparatory phase is apparently not stimulated by the object to which it is directed, for example not by the stick itself. If the ape sees a stick in a situation that does not require its use, except, for example, a roundabout way, it will not, of course, try to take hold of it, which means that the ape does not associate this phase of the activity with the stick but with the stick's objective relation to the fruit. The reaction to this [goal-oriented] relationship is nothing other than preparation for the next, second phase of the activity, i.e. the phase of realisation.
What is this second phase? It is already directed to[ward] the object [(i.e., biologically satisfying goal)] that immediately stimulates the animal, and is built up according to definite objective conditions, and consequently includes some operation or other that becomes a quite firm habit. [p. 192]
[In this]... highest stage of animal [psyche]... we thus observe a new complication in the structure of activity. The activity previously merged in a single process [of relatively immediate sensory or perceptual orientation toward biologically significant information] is now differentiated into two [intellectual] phases, one of preparation and one of accomplishment. Intellect arises for the first time, consequently, when preparation of the possibility to perform some operation or habit commences.
An essential attribute of two-phase [actions] is that new conditions no longer evoke simply trial movements in the animal but trials of previously developed ways or operations. How, for example, does a hen behave when driven out of an enclosure? It rushes blindly from side to side, trying to find a way out, i.e. simply increases its motor activity, until finally a chance movement leads to success. Higher animals behave differently in face of a difficulty. They also make trials, but these are not trials of separate movements but are primarily trials of various operations or modes of activity. Thus an ape, faced with a locked box, first tries the habitual operation of pressing on the lever; when that does not work, it tries to gnaw a corner of the box; then it employs a new method, to get into the box through the slit in the door. Then follows an attempt to gnaw off the lever, which is succeeded by an attempt to pull it off by its hand; finally, when that does not work, it employs the next method, to try and turn the box over [After Buytendijk, 1930].
This feature of ape's behaviour, which consists in their being able to solve [a novel]... problem in many ways, is most important evidence of their [repertoire of operations]... having ceased to [be] connected in [a] fixed way..., and [of their] not requiring the new problem.... to be directly similar to an earlier one.
Let us now consider [this] intellectual activity from the aspect of animal's reflection of their environment.
In its outward expression the first, ... phase of intellectual activity is directed to preparing for its second phase, i.e. is objectively governed by the next... [realisation phase]... itself. Does that mean, however, that the animal has its next operation in mind, i.e. that it is capable [p. 193] of imagining it [from the outset]? There is nothing to [unequivocally] justify such a supposition. The first phase corresponds to [both] the objective [perceptual] relation between things [and to the ongoing, active, consideration of the special situational relations of the particular problem itself]. This [situational] relation... must also be reflected by the animal... [for it to assess the success of the operations it is currently carrying out and for the selection of any other potential operation].....
...The transfer of an operation is now a transfer not only on the principle of the similarity of things (e.g. obstacles), with which the given operation was [formerly] associated, but also on the principle of the similarity of [situational] relations... to which [the animal] responds (e.g. branch-fruit). An animal [at the intellectual stage of psyche] now generalises the [perceptual] relations and [the ongoing situational] connections of things. Its [intellectual] generalisations are formed... exactly like the generalised, perceptual reflection of things, i.e. during the activity itself. [In other words, the preparatory activity elicited by the object of biological interest leads the psychic understanding of the animal. They understand the special situational relations better by way of doing. Activity leads understanding. Thus, by recognizing the goal-oriented aspects of the ape's two-phase actions without appealing to either an omnipresent, undifferentiated 'Gestalt principle,' or to the old arguments from 'preordained teleology,' the activity theory approach answers the methodological concerns of reductionist theory without falling prey to it.]
The origin and evolution of animal's intellect has its anatomical and physiological basis in a further development of the cerebral cortex and its functions. What are the main changes in the cortex observable in the higher stages of the evolution of the animal kingdom? The new thing that distinguishes the brain of higher mammals from that of lower animals is the relatively much greater place occupied by the frontal lobe, which is developed through differentiation of its prefrontal fields.
Study of the intellect of higher apes indicates that man's thinking has its real preparation in the animal kingdom, and that in this respect, too, there is no insuperable gulf between man and his animal ancestors. While noting the natural continuity in the evolution of the psyche in animals and man, however, one must not exaggerate their similarity in any way, as certain contemporary zoopsychologists do who try in their experiments with apes to demonstrate the alleged antiquity and naturalness of such 'intellectual behaviour' as working for pay and money exchange [See Wolfe, 1936].
Attempts to counterpose the intellectual behaviour of apes sharply to the behaviour of the higher mammals are also wrong. We now have many facts at our disposal [p. 194] that indicate that two-phase [actions] can be discovered in many higher animals including dogs, raccoons, and even cats (in the last named, which belong to the 'lurking' animals, it is true only in a very special expression).
Intellectual behaviour, which is proper to higher mammals, and which attains especially high development in apes, is thus the upper limit of the evolution of the [animal] psyche, after which the history of the evolution of a psyche of a quite different, new type begins, peculiar only to man, i.e. the history of the evolution of human consciousness.
4. The General Features of the Psyche of Animals
The prehistory of human consciousness is, as we have seen, the long, complex process of the evolution of animal's psyche..... [p. 195]....
The evolution of animal's reflection of their environment, however, also, as it were, lags behind the evolution of their activity. The simplest activity governed by the objective links of the affective properties and correlating the animal with a complex environment formed of things thus conditions the development of elementary sensations, which reflect only separate influences. The more complicated activity of vertebrates, determined by the physical relationships of things and situations, is linked with the [perceptual] reflection of whole things. Finally, when a 'phase of preparation' objectively determined by the possibilities of the animal's further action is differentiated at the stage of [animal] intellect, the form of the psyche is characterised by reflection of the physical relations of things and their physical situations. [p. 196]
The development of... psychic reflection is thus, as it were, ... shifted in relation to the evolution of the structure of animal's [leading] activity, so that there is never a direct correspondence between them.
Or rather, this correspondence can only exist as a moment marking a transition in evolution to the next, higher stage. Elimination of said disparity through the emergence of a new form of reflection opens up new possibilities of activity, which acquires an even higher structure, with the result that a disparity and contradiction again arises between them, but now already at a higher level....
In reviewing the evolution of the animal psyche we first of all stressed the differences that existed between its forms. Now we have to distinguish what these different forms have in common and what makes the activity of animals and their [p. 197] psyche qualitatively different from human activity and human consciousness.
The first difference... is that the former is instinctive, biological activity. In other words, animal activity can only be realised in relation to an object of vital, biological need or in relation to affective properties and things and their relationships (situations), that acquire the sense for animals of something that is connected with satisfying a certain biological need. Any change in animal activity therefore expresses in itself a change in the actual influence stimulating this activity, and not in the vital relationship itself that is realised by it. In ordinary experiments in forming a conditioned reflex in animals, for example, no new relation arises...; no new need develops in it, and if the animal now responds to the conditioned signal that is only because this signal now acts on it in the same way as an unconditioned stimulus. If we analyse any of an animal's diverse activities in general, we can always establish a certain biological relation that it realises, and consequently find the biological need underlying it.
Animal's activity thus always remains within the limits of their instinctive, biological relations with nature. That is a general law of animal activity.
In that connection the possibility of animal's psychic reflection of... reality... is also limited in principle. Because an animal enters into an interaction with... objects... affecting it, transferring its biological relations to them, it reflects only those aspects and properties of them that are connected with realising these relations.
Thus, whereas the figure of a triangle appears in man's consciousness irrespective of the actual relation to it, and is primarily characterised objectively, i.e. by the number of angles, etc., for an animal capable of distinguishing shape this figure is only distinguishable in so far as it has biological sense. A shape that is distinguished by an animal among several others will not be reflected by it apart from its appropriate biological relation. When an animal therefore has no instinctive relation with a given thing or a given [p. 198] affective property, and the thing has no connection with realisation of [its biological needs] the thing itself does not in that case exist, as it were, for the animal. In its activity it displays indifference to the influences concerned, which, although they could be an object of its perception, will never, however, become such in these conditions.... [p. 199].
Finally, we must note yet another essential feature of the psyche of animals that distinguishes it qualitatively from human consciousness. This is the fact that animal's relations to each other are the same in principle as their relations to other external objects, i.e. also belong exclusively to the realm of their... biological relations. That is connected with the fact that animals have no society [properly speaking]. We can observe the [individual] activity [i.e., operations or actions] of a few, [and] sometimes [the social joint action] of many, animals together, but we never observe [true] joint activity [proper] among them, i.e. joint in the [societal] sense of the word as we employ it when speaking of men's activity. For example, special observations of ants that are dragging a relatively big object along together -a twig of some sort or a big insect- indicate that the common, final path that their burden follows is not the result of... joint, organised [social] actions but is the result of the the mechanical addition of the efforts of individual ants, each one [operating on the object from the point of view of its merely sensory psyche] as if it were carrying the object independently. The same [, or rather, a similar sort of qualitative difference] is visible in the most highly organised animals, i.e. in apes. When several [p. 200] are faced with a problem requiring one box to be placed on another so that they can climb up on them and so reach a banana hanging high above..., each of the animals acts (as observations have shown) without considering the others. With such 'joint' action, therefore, a struggle for a box often develops, and a clash and fight between the animals, so that the 'structure' remains unbuilt, despite each ape separately knowing how, though not very dexterously, to pile one box on the other and clamber up...
In spite of these facts some writers consider that there is allegedly a division of labour among animals. For that they usually point to well-known examples from the life of bees, ants, and other 'social' animals. In fact, however, there is no real division of labour in all these cases,... as labour itself does not exist among them, i.e. a process that is by its very nature [societal-historical rather than merely a resultant of individual operations or even of social joint actions].
Although separate individuals among some animals do perform different functions in association, directly biological factors underlie this [de facto mechanical-collective, or social] difference of function. [In insects,] this is indicated both by the strictly defined, fixed character of the functions themselves (e.g. 'worker' bees build honeycomb, and later the queen bee lays eggs in them) and by the just as fixed character of their sequence (e.g. the consecutive change of functions of 'worker' bees). The division of functions in [truly cooperative] associations of higher animals, e.g. in a troop of apes, is of a more complex [social] character but it is determined in that case also by directly biological causes [i.e. goals]...
 The [biological] features of animal's relations with one another also determine the features of their 'speech' [or rather vocal communication]. As we know animal's communication is often expressed by one animal's affecting others by means of vocal sounds.... It applies, for example, to the signals given by sentry birds to the other members of the flock.
The [biological] features of animal's relations with one another also determine the features of their 'speech' [or rather vocal communication]. As we know animal's communication is often expressed by one animal's affecting others by means of vocal sounds.... It applies, for example, to the signals given by sentry birds to the other members of the flock.
In this case, however, do we have a process similar to the oral communication [i.e., meaningful language] of man? There is undoubtedly a certain similarity between them. But inwardly the processes are basically different. Man expresses a certain objective content in his speech and does not respond to speech addressed to him simply as sound stably associated with a definite [kind of] phenomenon, but rather to the reality reflected in the speech. We have quite another case in the vocal communication [p. 201] of animals. It can readily be demonstrated that [a bird] reacting to the voice of another animal, responds not to what the vocal signal objectively reflects but to the signal itself, which has acquired a certain biological sense for it.
If, for instance, we pick a chick up and restrain it by force it begins to struggle and cheep; its cheep attracts the hen..., which rushes in the direction of the sound and responds... by a special kind of clucking. This vocal behaviour of a chick and hen is outwardly similar to oral communication [even in humans]. But the process is in fact of quite a different nature.... That can easily be confirmed by the following experiment: if we put the restrained chick, which continues to cheep, under a thick glass cover that deadens the sounds, the hen, which can distinctly see the chick but no longer hears its cheeping, ceases to display any activity whatsoever in regard to it; the sight of the struggling chick in itself leaves [the hen] indifferent. The hen thus reacts not to what the chick's cheeping objectively means, in this case to a danger threatening it, but to the sound of the cheeping (see Fig. 30).
In principle the vocal behaviour of even more highly developed animals remains the same [(or at least similar)] in character, for example that of apes. As the findings of Yerkes and Learned [(1925)] indicate, it is impossible to teach apes real speech. [Put plainly, even the complex vocal signals of apes (in nature or otherwise) remains within the realm of traceable and relatively present biological-social significance. Their communication does not carry the sort of detached meaningful content or naming quality characteristic of the subject matter of human speech alone.]
It does not follow... that animal's vocal behaviour... is not associated with their psychic reflection of external objective reality, but as we have already said, the objects of their environments are inseparable for animals from their [own biological or social] relation... to these objects. An animal's expressive activity [whether vocal or otherwise] is therefore also [p. 202] never related to the objective thing itself [per se]. That is clearly seen from that fact that it repeats the very same vocal reaction when the biological sense of the given influences is identical for it, [even though] the objective... things may be completely different, rather than when the objects are identical in character. Birds that live in flocks, for example, have specific calls that warn the flock of danger. These cries are reproduced by the bird whenever it is frightened by something. At the same time it is quite irrelevant what [p. 203] precisely is affecting it in the case in point: one and the same call signals the appearance of a man, the appearance of a predator,or simply some sort of unusual noise. These calls are consequently linked with some phenomenon or other of reality, not by their objectively similar attributes, but simply by the similarity of the animal's instinctive relation to them. They are... associated with the animal's subjective states that arise in connection with these objects. In other words the animal cries mentioned lack a stable, objective, thing significance....
...[M]an's psyche is not only emancipated from those features that are common to all the stages of animal's psychic evolution that we have considered, and has not only acquired qualitatively new features, but (and this is the main point) the laws themselves that govern its evolution were altered with the transition to man. While the general laws governing the... psyche's evolution were those of biological evolution throughout the animal kingdom, with the transition to man... the psyche began to be governed by laws of socio-historical development.
[Section] II. The Origin of Human Consciousness [Human Intellect]
1. The Conditions of the Emergence of Consciousness
The transition to [human] consciousness is the beginning of a new, higher stage in the evolution of the psyche. In contrast to the psychic reflection peculiar to animals, conscious reflection [p. 204] is reflection of material reality in its separateness from the [individual] subject's... attitudes to it, i.e. reflection that distinguishes its objective stable properties.
...[W]hen I am aware, for example of this book, or even of my thought of [its theoretical relationship with other books], the book itself does not merge [(i.e., fuse)] in my consciousness with my [immediate tactile or perceptual] experience relative to the book, [n]or [does] the very idea of the book [become synonymous] with my [personal, individual or subjective] experience of this idea. [Rather, the book is reflected by me as both an "impending” object present to my individual experience and a culturally situated artifact -which was both materially produced by way of an extended collective system of human labor relations, and which is intended by the writer to remain (after his personal demise) as a tool of thought for others to use.]
The distinguishing of the reality reflected in man's consciousness has as another aspect the distinguishing of the world of inner experiences and the possibility of developing self-observation on that basis. [Humans can differentiate between objects, reflect upon relations between objects, and also (by the time of reaching adulthood) can form conscious judgments, plans, and decisions upon the basis of extended relations between relations (including societal-historical relations very distant in time and space from themselves).]
The task facing us is... to trace the conditions that give rise to this higher form of psyche...
The cause underlying the humanising of man's animal-like ancestors is the emergence of labour and the formation of human society on its basis. 'Labour,' Engels wrote, 'created man himself'. Labour also created man's consciousness.
The origin and development of labour, this first and basic condition for the existence of man, led to a change in his brain, his organs of external activity, and his sense organs, and their humanisation....
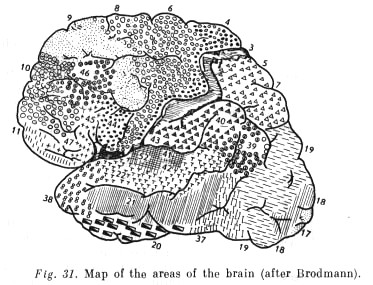 The main organ of man's labour, his hand, could only attain its perfection through the development of labour.... [p. 205]
The main organ of man's labour, his hand, could only attain its perfection through the development of labour.... [p. 205]
When we compare the maximum size of the simian skull and of the skull of primitive man, we find that the latter's brain is more than twice as big as that of the most highly developed modern species of ape (1400 cc and 600 cc).
The difference in the size of the brain of apes and man comes out even more sharply when we compare their weight; the difference is almost fourfold: the brain of an orangutan weighs 350 grams, and that of man 1400 grams.
Man's brain has a much more complicated, much more developed structure than the brain of the higher apes.
New areas of the cortex, not fully differentiated in the brain of apes, which later achieved full development in the brain of modern man, are already distinguishable in Neanderthal man, as casts of the inner surface of his skull indicate. Such are the areas numbered 44, 45, and 46 in Brodmann's system, in the frontal lobe, areas 39 and 40 in the parietal lobe, and areas 41 and 42 in the temporal lobe (see Fig. 31).
 It is very clear how new, specifically human features are reflected in the structure of the cerebral cortex when what we call the projection motor area is investigated (area 4 in Fig. 31). When various points in this area are carefully [p. 206] stimulated by an electrical current, ...the contractions of various groups of muscles [are stimulated]... Penfield expressed the results... in the form of the schematic... drawing... (see Fig. 32). It is clear from this figure what a relatively big surface is occupied in the human brain by the projections of such motor organs as the hands, and especially by the organs of speech (the muscles of the mouth,tongue, and ... larynx), whose functions are developed... intensively in the conditions of human society (labour, speech communication).
It is very clear how new, specifically human features are reflected in the structure of the cerebral cortex when what we call the projection motor area is investigated (area 4 in Fig. 31). When various points in this area are carefully [p. 206] stimulated by an electrical current, ...the contractions of various groups of muscles [are stimulated]... Penfield expressed the results... in the form of the schematic... drawing... (see Fig. 32). It is clear from this figure what a relatively big surface is occupied in the human brain by the projections of such motor organs as the hands, and especially by the organs of speech (the muscles of the mouth,tongue, and ... larynx), whose functions are developed... intensively in the conditions of human society (labour, speech communication).
Man's sense organs were also perfected thorough labour and in connection with the development of the brain. Like the organs of external activity they acquired qualitatively new features. The sense of touch became more exact, the humanised eye began to note more in things than the eye of the most farsighted birds, hearing developed capable of perceiving very fine differences and similarities in the sounds of human, articulated speech.
The development of the brain and sense organs in turn had a reverse effect on labour and speech, giving both 'an ever-renewed impulse to further development'.... [p. 207]....
The rise of labour was prepared [i.e., preadapted for] by the whole preceding course of evolution... The gradual transition to an upright posture, rudiments of which are distinctly observed even in the anthropoid apes...., and the forming... of specially mobile anterior extremities adapted to grasping objects, ... was due to the mode of life led by man's animal forebears -all created the physical preconditions for the possibility of performing complicated labour operations.
The labour process was also.... only possible ... to emerge in animals that lived in whole groups and which had sufficiently developed [social] forms of joint life, although those forms were... very remote even from the most primitive forms of human [societies]... The interesting research ... by Voitonis and Tikh at the Sukhumi nursery.... has shown there is already an established system of mutual relations within a troop of apes and a peculiar hierarchy with a correspondingly... complex system of [social] intercourse....
An essential precondition of labour, finally, was also the existence among the highest members of the animal kingdom, as we have seen, of very developed forms of the psychic reflection of reality [(i.e., those necessary for the successful performance of two-phase problem solving actions)].
In the aggregate all these [preadaptions]... constituted the main conditions by which, in the course of subsequent evolution, labour, and human society based on labour, could arise.
What is this specific human activity that we call labour?
Labour is the process that links man with nature, the process of man's action on nature. [p. 208]....
The two following features are above all typical of labour. The first is the use and making of tools....
The second feature... is that it is performed in conditions of joint, collective activity, so that man functions in the process not only in a certain relationship with nature but also to other people, members of a given society. Only through a relation with other people does man relate to nature itself, which means that labour [as an emergent societal construct] appears from the very beginning as a process mediated by [(i.e., carried out via the use of)] tools (in the broad sense) and at the same time mediated socially.
Man's [characteristically societal-historical] use of tools also has a natural history of its [social] preparation. There are already the rudiments of tool [use] among certain animals as we know, in the form of the use of external means with whose aid they perform separate [i.e., relatively individual] operations [or actions] (e.g. an ape's use of a stick). These external means -animal's 'tools'- differ qualitatively, however, from man's true tools, [which function as] the implements of [collective] labour.
The difference between them by no means consists solely in the animal's using his 'tool' more seldom than [did hominids or] primitive men. Even less does their difference consist in differences simply in their outward form [i.e., the mere structural sophistication of the tool itself]. We can only bring out the real difference between human tools and animal's 'tools' by making an objective review of the [specific forms of] activity... in which they are [or were] included.
However complex animal's 'tool' [use seems], it never has the social [i.e., truly collective] character [of its human counterpart]... and does not itself govern a relationship of community among the individuals performing it.... [p. 209]....
In contradistinction, human labour is social activity from the beginning, based on the [necessary] cooperation of individuals, assuming a technical division... of labour functions; labour consequently is a [special, societally contextualized] process of action on nature linking together its [social] participants, and mediating their contact....
It is sufficient, in order to comprehend the concrete significance of this fact for the the development of the human psyche, to analyse how the structure of [that] activity is altered when it is performed in conditions of collective labour.
Already at the earliest time in the evolution of human society, a division of the previously single process of ...[tool] production... arose.... In the course of subsequent evolution it took shape... as a primitive technical division of labour.
It now fell to the lot of some individuals, for example, to maintain the fire [i.e., a tool that no animal uses] and to cook food on it, and of others to procure the food itself. Some of those taking part in the collective hunt fulfilled the function of pursuing game, others the function of waiting for it in ambush and attacking it.
[Although lions and apes also carry out similar joint actions during their pursuit of live prey (i.e., thereby providing them with relatively direct satisfaction of their biological needs) the allocation of the acquired game is carried out according to an immanently traceable biological dominance hierarchy or through what might be described as socially organized 'scrounging'). It was with the adoption of mediating tools in our ancestral hominids (e.g., for killing, preparing, cooking, and then storing acquired game), however, that a new predominance and pervasiveness of such socially situated two-phase actions was stretched out over both time and space.]
This [emerging pattern of collective social-societal relations] led to a decisive, radical change in the very structure of the [psychical] activity of the individuals taking part in the labour process.... [p. 210]....
Let us now examine the fundamental structure of the individual's activity in the condition of a collective labour process from this standpoint. When a member of a group performs his labour activity he also does it to satisfy one of his needs. A beater, for example, taking part in a primaeval collective hunt, was stimulated by a need for food or, perhaps, a need for clothing, which the skin of [a] dead animal would meet for him. [This, properly speaking, would be his motive for participating in the hunt.] At what [goal], however, was his [role in the hunt] directly aimed? It may have been [aimed]... at frightening a herd of animals and sending them toward other hunters, hiding in ambush. That, properly speaking, is what should be the result of the activity of this man. And the activity of this individual member of the hunt ends with that. The rest is completed by the other members. [The result of his particular actions], i.e. the frightening of game, etc. ... does not in itself... lead to satisfaction of the beater's need for food, or the skin of the animal. [The goals of his actions]... did not, ... coincide with what stimulated them, i.e. did not coincide with the motive of his activity; the two were divided from one another... We can say, for example that the beater's activity is the hunt, and the frightening of game his action.
.... The product of [this sort of joint, collective process of acting on nature] which meets the need of the group, also leads to satisfaction of the needs of the separate individual as well, although he himself may not perform the final operations (e.g. the direct attack on the game and the killing of it)... Genetically (i.e. in its origin) the separation [p. 211] of the [goals of an individual's actions] and [the overlying] motive of individual activity is a result of the [gradual, evolutionarily selected,] disarticulating [i.e., the stretching out over time or space] of the separate ["preparation and realisation"] operations [already noted (in their preadaptive rudimentary forms) to be characteristic of the two-phase actions of higher animals], .... for example among apes.... Experiments [with animals] have shown, for example, that a forced break... [within] one of its phases makes it possible to postpone animal's subsequent reaction only a very little, while a break between phases gives a postponement ten or even a hundred times as long in the same animal (Zaporozhets' experiments).
In spite of the existence of an undoubted [preadaptive] link between the two-phase intelligent [actions] of higher animals and the [two-phase] activity of the individual man taking part in a collective labour process..., there is a vast difference between them, a difference that is... reflected in the psyche of the acting individuals.
The peculiarity of animal's two-phase intelligent [actions] consists, as we have seen, in the link between its two (or even several) phases being determined by [relatively immediate] physical, material links and relations (spatial, temporal, and mechanical). In animal's natural conditions of existence, furthermore, these are always natural [i.e., bio-social] links and relations....
When an animal, in taking a roundabout method, first [p. 212] turns away from bait and only then seizes it, this complicated [two-phase action toward a goal] is governed by the spatial relations of the situation perceived by the animal; the first part of the path... presents the animal of necessity with the possibility of performing the second phase.
The form of [typical two-phase] human activity... has a decidedly different objective basis.
....In that case.... [the “disarticulated” connection between a goal and a motive]... is realised through the activity of other people, which means that it is the activity of other people that constitutes the objective basis of the specific structure of [a] human individual's activity, i.e. ... the connection between the motive [of an activity] and the [goal] of an action reflects objective social [and historical]... relations rather than [merely bio-social] ones.
The complex [individual actions] of higher animals governed by natural material connections and [bio-social] relations is thus converted in man into activity that is governed by connections and relations that are primordially social. That [novel societal context] also constitutes the direct reason why a specifically human form of reflection of reality, human consciousness, arises.
The [routine, practical] separation of [individual action and collective activities within human society] necessarily presupposes the possibility of the active subject's psychic reflection of the relation between the [general] objective motive and the [particular] object [i.e., goal] of the action. Otherwise the [particular] action... lacks any sense for its subject.... [p. 213]....
Thus, together with the birth of action, this main 'unit' in human activity, there also arises the main unit, social in nature, of the human psyche, i.e. the rational meaning for man of that which his activity is directed to....
Basic "Structure” and terms used with regard to Human Activity:
Activity (in human beings) is governed by its motive/motives: The man is engaged in a communal hunt because he wants to feed his family.
Actions are governed by their goals: The man performs the role of “beater” (the goal being to scare the prey away from himself and toward the other members of the hunting party).
Operations are governed by the conditions of the hunt. How he carries out the various tasks involved in his role will depend upon the terrain, kind of game-animal sought, wind direction, the weather, the season of the year, etc.
Thanks to Paul F. Ballantyne, Ph.D., April, 5, 2005, pballan@comnet.ca
{kind=link}